Out of the Blue (Ocean)
The adventures of a marine ecologist!


Ecology Explained: The Intermediate Disturbance Hypothesis
The Intermediate Disturbance Hypothesis is one of the staples of ecology, especially marine ecology. The Intermediate Disturbance Hypothesis was first proposed by Connell, a well-known intertidal and general ecologist, in 1978 (See his article “Diversity in Tropical Rain Forests and Coral Reefs”). But what is it exactly? Let me explain!
Let’s start with this handy diagram from Wikipedia.
As you can see, we have Level of Disturbance on the x-axis. That simply describes the level of disturbance present in the system. It could be any sort of disturbance…fires, hurricanes, waves, human or animal trampling, wind, sun, and so on. The level of disturbance increases from left to right. So, the area marked with I has less disturbance than area II.
Species Diversity on the y-axis is more of a general term. Sometimes this is “species richness,” which is a pure count of species present in the ecosystem. Sometimes this is actually referring to “species diversity,” which takes the number of species from species richness and combines it with how the species are distributed in the system. But that’s a subject for another blog post! All you need to know now is that species diversity in the system increases from the bottom to the top.
Now let’s look at those areas marked by Roman numerals.
Area I is first up. In that section, we have a low amount of disturbance, which results in an okay amount of diversity. Why is that?
Ecosystems typically have a successional pathway–a pattern across time of species that are present. Think about a forest directly after a forest fire. Early-regrowth forests are going to look totally different from established forests! Those established forests will most likely have one or more competitive dominants–the species that compete the best. That’s great for the competitive dominants because they do well in those established systems, but it’s not so good for diversity. The competitive dominants compete so well that there isn’t much room for other species.
(Sometimes you will see similar graphs that are a simple bell curve with Area I showing the same low, low diversity that Area III exhibits. This can also happen–if there is no disturbance in the system at all, the species diversity is going to be extremely low. But, from my perception, it’s probably not going to get as bad as Area III. It’s all up to interpretation.)
Area II looks great! There is an intermediate amount of disturbance and the maximum amount of diversity. In Area II, there is enough disturbance in the system to stop the competitive dominants from over-dominating. Organisms earlier in the successional pathway that are poorer competitors (but still play an important role) are able to survive. This results in the maximum amount of species diversity! Hence the name “Intermediate Disturbance Hypothesis!” This area is experiencing an intermediate amount of disturbance, but not enough to push it to Area III.
Area III is not a good place to be. Area III exhibits a very high amount of disturbance and a low amount of diversity. It’s pretty easy to understand why! Think of a coral reef that’s constantly being battered by huge hurricanes or a forest that has repeated, huge fires. The succession pathway barely even gets a chance to begin before another huge disturbance sweeps through. This will result in very low diversity.
So, the Intermediate Disturbance Hypothesis shows us that with an intermediate level of disturbance we can expect a high amount of diversity. With low and high levels of disturbance, not so much!
Do you have a lingering question? Ask it in the comments section and I will be happy to help as best I can!
Share this:
Leave a comment cancel reply.
- Already have a WordPress.com account? Log in now.
- Subscribe Subscribed
- Copy shortlink
- Report this content
- View post in Reader
- Manage subscriptions
- Collapse this bar
- The Open University
- Guest user / Sign out
- Study with The Open University
My OpenLearn Profile
Personalise your OpenLearn profile, save your favourite content and get recognition for your learning
About this free course
Become an ou student, download this course, share this free course.

Start this free course now. Just create an account and sign in. Enrol and complete the course for a free statement of participation or digital badge if available.
4.2 The intermediate disturbance hypothesis
The realisation that the frequency of disturbance can influence community structure led to the formulation of the intermediate disturbance hypothesis (IDH) (Connell, 1978; Figure 22 below). The IDH proposes that species diversity is generally maximised if disturbance is neither too rare nor too frequent because species that thrive at both early and late successional stages can coexist.

A hypothetical line graph that shows how species diversity changes with frequency of disturbance. The horizontal axis is labeled ‘frequency of disturbance’ and the vertical axis is labeled ‘species diversity’. There are no quantitative markings on the vertical axis. On the horizontal axis, the extreme left-hand side is labeled ‘low’ and the extreme right hand side ‘high’. The line of the graph starts on the bottom left, increases almost linearly, reaches a peak and then decreases linearly to reach a low value again. So, it is an inverted U-shape. Underneath the graph is a series of three pictures of vegetation. The one on the left which is aligned with the left-hand side of the graph, shows a landscape with mostly grass interspersed with a few small trees. The central picture, which is aligned with the peak on the graph shows a mixture of low vegetation such as grass and shrubs, small trees and larger trees. The third picture which is aligned with the right-hand side of the graph shows mostly large trees. The graph and the pictures together illustrate that at a low frequency of disturbance (LHS of the graph, LH picture), species diversity is low; at an intermediate frequency of disturbance (peak on graph, central picture) diversity is high and reaches a peak; and at a high frequency of disturbance (RHS of graph, RH picture) diversity is once again low.
Connell’s original paper has received more than 4000 citations and is still referenced in important scientific papers. Many studies have empirically validated the IDH, particularly for marine systems. However, there are also an increasing number that show little support and this has led to a great deal of controversy regarding its validity in explaining the relationship between disturbance and species diversity. For example, it can be argued that tropical forests show high diversity even though natural disturbance is minimal.
Whether or not the IDH holds appears to depend, to some degree, on scale (whether on a local or geographical scale) and the type of disturbance. For example, at a small local scale species diversity is often maximised at a high frequency of fire rather than at an intermediate or low frequency.
Give an example where frequent fire is necessary to maintain species diversity.
The Fynbos in the Cape region of South Africa.
However on a larger scale, disturbances of intermediate frequency and/or intensity may generate diversity. For example, a fire of intermediate or mixed severity could increase diversity by generating spatial heterogeneity within a landscape.
This is because a mixed severity fire will result in a complex of patches in a landscape. Patches differ in severity of burn and include unburned patches, low severity burn patches, moderate severity burn patches where perhaps one-third to two-thirds of the vegetation is killed, and high severity burn patches where almost all the vegetation is killed (Figure 23). In this case, the IDH may not hold on a local scale (within patches) because species richness within a small area may decline (for example, in severely burnt areas). However, on a larger landscape scale a fire of intermediate severity may increase species diversity. This is because areas or patches that differ in burn severity, and that are at different stages of post-fire change, would sustain more species with different disturbance sensitivities.

A greyscale image that shows the burn severity. White patches show low severity burn, grey patches show moderate severity burn, and black patches show high severity burn.
Although the IDH is controversial, the idea that landscapes with greater heterogeneity in size, age, and burn severity of post-fire patches support a greater diversity of species, has led some researchers to propose that a diversity of fire regimes across a landscape (named pyrodiversity ) is necessary to maintain biodiversity.
Intermediate Disturbance Hypothesis in Ecology: A Literature Review
- Related Documents
Testing the intermediate disturbance hypothesis on species diversity in herbaceous plant communities along a human trampling gradient using a 4-year experiment in an old-field
Diversity–disturbance relationships: frequency and intensity interact.
An influential ecological theory, the intermediate disturbance hypothesis (IDH), predicts that intermediate levels of disturbance will maximize species diversity. Empirical studies, however, have described a wide variety of diversity–disturbance relationships (DDRs). Using experimental populations of microbes, we show that the form of the DDR depends on an interaction between disturbance frequency and intensity. We find that diversity shows a monotonically increasing, unimodal or flat relationship with disturbance, depending on the values of the disturbance aspects considered. These results confirm recent theoretical predictions, and potentially reconcile the conflicting body of empirical evidence on DDRs.
Short-term response of estuarine sandflat trophodynamics to pulse anthropogenic physical disturbance: Support for the Intermediate Disturbance Hypothesis
The intermediate disturbance hypothesis explains arthropod beta-diversity responses to roads that cut through natural forests, intermediate disturbance hypothesis in phytoplankton ecology, a phytoplankton community in a temperate reservoir in new south wales, australia: relationships between similarity and diversity indices and measures of hydrological disturbance.
Temporal changes in diversity and similarity of a phytoplankton community were investigated in relation to external hydrological disturbance in the Ben Chifley reservoir from September 1998 to January 2002. Species richness varied by a factor of 4–5 at each of three sites studied during the period (n = 53 at each site). Species diversity (measured using Simpson’s D and Shannon–Wiener’s H, based on primarily genus or species number and cell densities) varied by a factor of 8–10, whereas similarity between two consecutive sampling dates (measured using Hurlbert’s index and Pinkham and Pearson’s B) varied by a factor of 10–46. When diversity was measured with H, it had an approximate quadratic (convex) relationship with similarity, as measured with Hurlbert’s index. However, diversity was seldom related to external hydrological disturbance (measured as intensity and variability of daily inflow rates between two consecutive sampling dates). Similarity was significantly and negatively related to disturbance variability. These results suggest that the mechanisms that regulate diversity and similarity may differ from each other, and question the usefulness of observed approximate quadratic relationships between similarity and diversity indices when assessing the effect of disturbance on diversity. Such relationships may therefore not provide support for Connell’s (1978) intermediate disturbance hypothesis.
Hurricanes and coral reefs: The intermediate disturbance hypothesis revisited
Theory of marine communities: the intermediate disturbance hypothesis, effects of forest roads on hemipteran diversity in mt. gariwang, korea test of intermediate disturbance hypothesis, hypertrophic phytoplankton and the intermediate disturbance hypothesis, export citation format, share document.

- school Campus Bookshelves
- menu_book Bookshelves
- perm_media Learning Objects
- login Login
- how_to_reg Request Instructor Account
- hub Instructor Commons
Margin Size
- Download Page (PDF)
- Download Full Book (PDF)
- Periodic Table
- Physics Constants
- Scientific Calculator
- Reference & Cite
- Tools expand_more
- Readability
selected template will load here
This action is not available.
18.2: What are the Effects of Disturbance?
- Last updated
- Save as PDF
- Page ID 78508
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
Changes in community structure and composition over time are induced by environmental disturbances such as volcanoes, earthquakes, storms, fires, and climate change. Communities with a stable structure are said to be at equilibrium. Following a disturbance, the community may or may not return to the equilibrium state. In primary succession, newly exposed or newly formed land is colonized by living things; in secondary succession, part of an ecosystem is disturbed and remnants of the previous community remain. Thus, disturbance can initiate successional change.
Species that are well adapted for exploiting disturbance sites are referred to as pioneers or early successional species. These shade-intolerant species are able to photosynthesize at high rates, produce a lot of offspring, and grow and mature quickly. Their fast growth is usually balanced by short life spans. Furthermore, although these species often dominate immediately following a disturbance, they are unable to compete with shade-tolerant species later on and are replaced by these species through succession. However these shifts may not reflect the progressive entry to the community of the taller long-lived forms, but instead, the gradual emergence and dominance of species that may have been present, but inconspicuous directly after the disturbance. 1 Disturbances have also been shown to be important facilitators of non-native plant invasions. 2
While plants must deal directly with disturbances because of their lack of mobility, many animals are mobile and thus are not as immediately affected by disturbance. For example, some animals could successfully evade the initial destruction of a forest fire, but can later return to the burned area and thrive on new growth on the forest floor. Disturbed communities (such as a forest after a fire) often support a wider variety of plants compared to pre-disturbance vegetation. The plants in turn support a variety of wildlife, temporarily increasing biological diversity in the forest. 3
Intermediate Disturbance Hypothesis
The intermediate disturbance hypothesis (IDH) suggests that local species diversity is maximized when ecological disturbance is neither too rare nor too frequent. At low levels of disturbance, more competitive organisms will push subordinate species to extinction and dominate the ecosystem. 4 At high levels of disturbance, due to frequent forest fires or human impacts like deforestation, all species are at risk of going extinct. According to intermediate disturbance hypothesis theory, at intermediate levels of disturbance, diversity is thus maximized because species that thrive at both early and late successional stages can coexist. Intermediate disturbance hypothesisis a nonequilibrium model used to describe the relationship between disturbance and species diversity.
The intermediate disturbance hypothesis is based on the following premises:
- ecological disturbances have major effects on species richness within the area of disturbance,
- interspecific competition results from one species driving a competitor to extinction and becoming dominant in the ecosystem, and
- moderate ecological scale disturbances prevent interspecific competition. 5,6,7
Figure \(\PageIndex{1}\): I describes how, at low levels of ecological disturbance species richness decreases as competitive exclusion increases; II shows that at intermediate levels of disturbance, diversity is maximized because species that thrive at both early and late successional stages can coexist; III shows that at high levels of disturbance species richness is decreased due to an increase in species movement. " Intermediate Disturbance Hypothesis Graph " by Sciencerelatedusername is licensed under CC BY-SA 4.0.
Disturbances act to disrupt stable ecosystems and clear species' habitat. As a result, disturbances lead to species movement into the newly cleared area (secondary succession). 5 Once an area is cleared there is a progressive increase in species richness and competition between species takes place. Once the conditions that create a disturbance are gone, and competition between species in the formerly disturbed area increases, species richness decreases as competitive exclusion increases. 8
"Gause's Law", also known as competitive exclusion, explains how species that compete for the same resources cannot coexist in the same niche. 6 Each species handles change from a disturbance differently; therefore, intermediate disturbance hypothesis can be described as both "broad in description and rich in detail". 5 The broad intermediate disturbance hypothesis model can be broken down into smaller divisions which include spatial within-patch scales, spatial between-patch scales, and purely temporal models. Each subdivision within this theory generates similar explanations for the coexistence of species with habitat disturbance. Joseph H. Connell 9 proposed that relatively low disturbance leads to decreased diversity and high disturbance causes an increase in species movement. These proposed relationships lead to the hypothesis that intermediate disturbance levels would be the optimal amount of disorder within an ecosystem.
Another way of thinking about the intermediate disturbance hypothesis requires that we consider the types of organisms that could specialize in areas with different levels of disturbance. K-selected species generally demonstrate more competitive traits. Their primary investment of resources is directed towards growth, causing them to dominate stable ecosystems over a long period of time. In contrast, r-selected species colonize open areas quickly and can dominate landscapes that have been recently cleared by disturbance. 7 These characteristics attribute to the species that thrive in habitats with higher and lower amounts of disturbance. Based on the contradictory characteristics of both of these examples, areas of occasional disturbance allow both r and K species to flourish in the same area. If K-selected and r-selected species can live in the same region, species richness can reach its maximum.
Several alternative hypotheses to the intermediate disturbance hypothesis have been proposed 10 . One alternative hypothesis states that the species diversity in a disturbance-mediated coexistence between species is maximized by the presence of a disturbance regime resembling the historic processes. This is because species generally adapt to the level of disturbance in their ecosystem through evolution (whether disturbance is of high, intermediate or low level). In addition, many species (e.g. ruderal plants and fire-adapted species) even depend on a specific disturbance in ecosystems where it often occurs.
- Nobel, I. The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances.
- Lembrechts, J.J., Pauchard, A., Lenoir, J., Nuñez, M.A., Geron, C., Ven, A., Bravo-Monasterio, P., Teneb, E., Nijs, I., & Milbau, A. (2016). Disturbance is the key to plant invasions in cold environments . Proceedings of the National Academy of Sciences, 113 (49), pp. 14061–14066. doi : 10.1073/pnas.1608980113 . ISSN 0027-8424 . PMC 5150417 . PMID 27872292 .
- Pringle, L. (1979). Natural fire: Its ecology in forests. William Morrow and Company, New York, pp . 27-29.
- Dial, R., & Roughgarden, J. (1988). Theory of marine communities: The intermediate disturbance hypothesis. Ecology, 79 (4), pp. 1412–1424. doi : 10.1890/0012-9658(1998)079[1412:TOMCTI]2.0.CO;2
- Wilkinson, D.M. (1999). The disturbing history of intermediate disturbance. Oikos, 84 (1), pp. 145–7.
- Kricher, J.C. (2011). Tropical Ecology. New Jersey, Princeton: Princeton University Press .
- Catford, J.A., Daehler, C.C., Murphy, H.T., Sheppard, A.W., Hardesty, B.D., Westcott, D.A., Rejmánek, M., Bellingham, P.J., et al. (2012). The intermediate disturbance hypothesis and plant invasions: Implications for species richness and management. Perspectives in Plant Ecology, Evolution and Systematics, 14 (3), pp. 231–41. doi : 10.1016/j.ppees.2011.12.002
- Vandermeer, J., Boucher, D., Perfecto, I., & de la Cerda, I.G. (1996). A theory of disturbance and species diversity: Evidence from Nicaragua after Hurricane Joan. Biotropica, 28 (4), pp. 600–13. doi : 10.2307/2389100 . JSTOR 2389100
- Connell, J.H. (1978). Diversity in tropical rain forests and coral reefs. Science, 199 (4335), pp. 1302–10. Bibcode : 1978Sci...199.1302C
- Hall, A.R., Miller, A.D., Leggett, H.C., Roxburgh, S.H., Buckling, A., & Shea, K. (2012). Diversity-disturbance relationships: Frequency and intensity interact . Biology Letters, 8 (5), pp. 768–71. doi : 10.1098/rsbl.2012.0282
Contributors and Attributions
Modified by Castilleja Olmsted (University of Pittsburgh) and Kyle Whittinghill (University of Vermont) from the following sources:
- Connie Rye (East Mississippi Community College), Robert Wise (University of Wisconsin, Oshkosh), Vladimir Jurukovski (Suffolk County Community College), Jean DeSaix (University of North Carolina at Chapel Hill), Jung Choi (Georgia Institute of Technology), Yael Avissar (Rhode Island College) among other contributing authors. Original content by OpenStax (CC BY 4.0; Download for free at http://cnx.org/contents/[email protected] ).
- https://en.wikipedia.org/wiki/Interm...nce_hypothesis
- https://en.wikipedia.org/wiki/Disturbance_(ecology)

Want to create or adapt books like this? Learn more about how Pressbooks supports open publishing practices.
17 Chapter 17: Disturbance and Succession
Lisa Limeri
Learning Objectives
By the end of this section, students will be able to…
- Describe primary and secondary succession and the three models of succession: facilitation, inhibition, and tolerance.
- Describe the relationship between disturbance regime (frequency and intensity) and biodiversity in a community.
Ecological Disturbance
Changes in community structure and composition over time are induced by environmental disturbances such as volcanic eruptions, earthquakes, storms, fires, and climate change. Communities with a stable structure are said to be at equilibrium. Following a disturbance, the community may or may not return to the equilibrium state. In primary succession , newly exposed or newly formed land is colonized by living things; in secondary succession , part of an ecosystem is disturbed and remnants of the previous community remain. Thus, disturbance can initiate successional change.
Species that are well adapted for exploiting disturbance sites are referred to as pioneers or early successional species. In forests, these shade-intolerant species are able to photosynthesize at high rates and as a result grow quickly. Their fast growth is usually balanced by short life spans. Furthermore, although these species often dominate immediately following a disturbance, they are unable to compete with shade-tolerant species later on and are replaced by these species through succession. However these shifts may not reflect the progressive entry to the community of the taller long-lived forms, but instead, the gradual emergence and dominance of species that may have been present, but inconspicuous directly after the disturbance. Disturbances have also been shown to be important facilitators of non-native plant invasions.
While plants must deal directly with disturbances because of their lack of mobility, many animals are mobile and thus are not as immediately affected by disturbance. For example, some animals could successfully evade the initial destruction of a forest fire, but can later return to the burned area and thrive on new growth on the forest floor. Disturbed communities (such as a forest after a fire) often support a wider variety of plants compared to pre-disturbance vegetation. The plants in turn support a variety of wildlife, temporarily increasing biological diversity in the forest.
Reading Question #1
Which of the following terms describes the first species to colonize newly-formed land, such as the result of a volcanic eruption?
A. Climax community B. Keystone species C. Foundation species D. Pioneer species
Intermediate Disturbance Hypothesis
The intermediate disturbance hypothesis (IDH) suggests that local species diversity is maximized when ecological disturbances are neither too rare nor too frequent (Figure 17.1). At low levels of disturbance, more competitive organisms will push subordinate species to extinction and dominate the ecosystem. At high levels of disturbance, due to frequent forest fires or human impacts like deforestation, all species are at risk of going extinct. According to IDH, at intermediate levels of disturbance, diversity is thus maximized because species that thrive at both early and late successional stages can coexist. IDH is a non-equilibrium model used to describe the relationship between disturbance and species diversity.
IDH is based on the following premises:
- ecological disturbances have major effects on species richness (the total number of different species) within the area of disturbance,
- interspecific competition results in one species driving a competitor to extinction and becoming dominant in the ecosystem, and
- moderate ecological scale disturbances prevent interspecific competition.
Disturbances act to disrupt stable ecosystems and clear species’ habitat. As a result, disturbances lead to species movement into the newly cleared area (secondary succession). Once an area is cleared there is a progressive increase in species richness and competition between species takes place. Once the conditions that create a disturbance are gone, and competition between species in the formerly disturbed area increases, species richness decreases as competitive exclusion increases.
Reading Question #2
When will biodiversity be highest?
A. Where there is no disturbance in the ecosystem B. When there is an intermediate level of disturbance in the ecosystem C. When there is a high level of disturbance in the ecosystem D. When there are variable levels of disturbance in the ecosystem
Ecological Succession
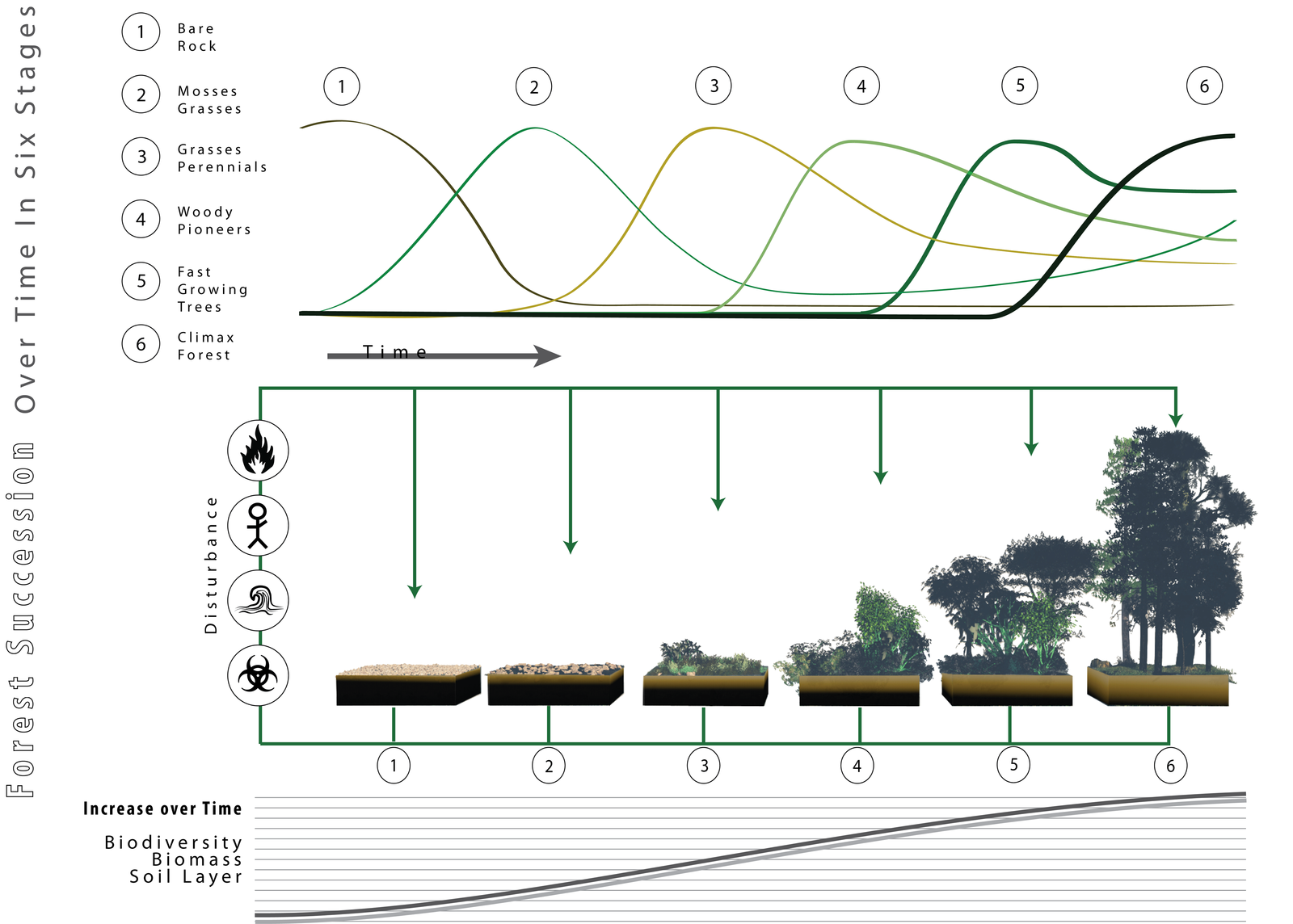
Communities with a stable structure are said to be at equilibrium. Following a disturbance, the community may or may not return to the equilibrium state. Succession describes the sequential appearance and disappearance of species in a community over time following disturbance. There are two types of ecological succession: primary and secondary. In primary succession , newly exposed or newly formed land is colonized by living things; in secondary succession , part of an ecosystem is disturbed and remnants of the previous community remain.
Primary Succession
Primary succession occurs when new land is formed or rock is exposed: for example, following the eruption of volcanoes, such as those on the Big Island of Hawaii. As lava flows into the ocean, new land is continually being formed. On the Big Island, approximately 32 acres of land are added each year. First, weathering and other natural forces break down the substrate enough for the establishment of pioneer species such as hearty plants and lichens with few soil requirements (Figure 17.2). These species help to further break down the mineral-rich lava into the soil where other, less hardy species will grow and eventually replace the pioneer species. In addition, as these early species grow and die, they add to an ever-growing layer of decomposing organic material and contribute to soil formation. Over time the area will reach an equilibrium state, with a set of organisms quite different from the pioneer species.

Secondary succession
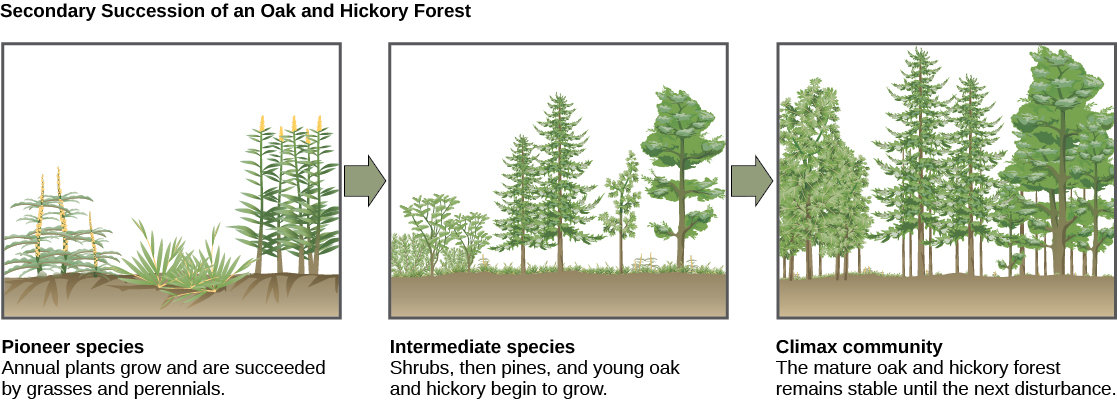
A classic example of secondary succession occurs in oak and hickory forests cleared by wildfire (Figure 17.3). Wildfires will burn most vegetation and kill those animals unable to flee the area. Their nutrients, however, are returned to the ground in the form of ash. Thus, even when areas are devoid of life due to severe fires, the area will soon be ready for new life to take hold.
Before the fire, the vegetation was dominated by tall trees with access to the major plant energy resource: sunlight. Their height gave them access to sunlight while also shading the ground and other low-lying species. After the fire, though, these trees are no longer dominant. Thus, the first plants to grow back are usually annual plants followed within a few years by quickly growing and spreading grasses and other pioneer species. Due to, at least in part, changes in the environment brought on by the growth of the grasses and other species, over many years, shrubs will emerge along with small pine, oak, and hickory trees. These organisms are called intermediate species. Eventually, over 150 years, the forest will reach its equilibrium point where species composition is no longer changing and resembles the community before the fire. This equilibrium state is referred to as the climax community, which will remain stable until the next disturbance.
Climatic factors may be very important, but on a much longer time-scale than any other. Changes in temperature and rainfall patterns will promote changes in communities. As the climate warmed at the end of each ice age, great successional changes took place. The tundra vegetation and bare glacial till deposits underwent succession to mixed deciduous forest. The greenhouse effect resulting in increase in temperature is likely to bring profound Allogenic changes in the next century. Geological and climatic catastrophes such as volcanic eruptions, earthquakes, avalanches, meteors, floods, fires, and high wind also bring allogenic changes.
In general, communities in early succession will be dominated by fast-growing, well-dispersed species (opportunist, fugitive, or r-selected life histories). As succession proceeds, these species will tend to be replaced by more competitive (K-selected) species. Trends in ecosystem and community properties in succession have been suggested, but few appear to be general. For example, species diversity almost necessarily increases during early succession as new species arrive, but may decline in later succession as competition eliminates opportunistic species and leads to dominance by locally superior competitors.
Reading Question #3
What is the difference between primary and secondary succession?
A. Primary succession always occurs before secondary succession B. Primary succession occurs more often than secondary succession C. In primary succession a brand new community is founded whereas in secondary succession the community is influenced by the pre-existing community D. In primary succession, only pioneer species can establish whereas in secondary succession only climax species can establish.
Climax Communities
Ecological succession was formerly seen as having a stable end-stage called the climax community, shaped primarily by the local climate (Figure 17.4). This idea has been largely abandoned by modern ecologists in favor of non-equilibrium ideas of ecosystem dynamics. Most natural ecosystems experience disturbance at a rate that makes a “climax” community unattainable. Climate change often occurs at a rate and frequency sufficient to prevent arrival at a climax state. Additions to available species pools through range expansions and introductions can also continually reshape communities.
Models of Successional Change
Joseph Connell and Ralph Slatyer further developed the understanding of successional mechanisms in their 1977 paper and proposed that there were 3 main modes of successional development: Facilitation , Tolerance , and Inhibition .
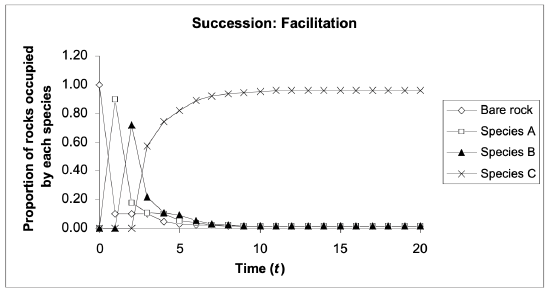
Facilitation model
The facilitation model is based on the assumption that only particular species with qualities ideal for “early succession” can colonize the newly exposed landforms after an ecological disturbance.
- These “colonizing” qualities include: highly effective methods of dispersal, the ability to remain dormant for long periods of time, and a rapid growth rate. However, the pioneer species are often subsequently less successful once an area has been heavily populated by surrounding species due to increased shade, litter or concentrated roots in the soil, etc.
- Thus, the presence of early successional species often changes the environment so that the habitat is less hospitable for the original species’ own ecological demands and facilitates the invasion of later-successional species.
The facilitation model suggests that the presence of an initial species aids and increases the probability of the growth of a second species (Fig 17.5). For example, the presence of alder plants aids the growth of willow and poplar seedlings in an Alaskan floodplain. Alder roots contain nitrogen-fixing bacteria, which greatly increase the amount of inorganic nitrogen present in soils. This increased availability of nitrogen aids the growth of both willow and poplar seedlings in areas without other competition. Eventually, however, willow and poplar grow more rapidly than alder, leading to a reduction in the abundance of the pioneer species, and eventually, spruce becomes a later-succession species, due to its increased ability (over alder) to grow in shaded areas.
Another case of facilitation comes from the colonization of lakeshore sand dunes. Adjacent pioneer plants colonize the otherwise moving sands and alter the environmental constraints of the sandy environment to better suit other plant species, which can then allow for soil binding to take place. The giant saguaro cactus, in this respect, can only survive in the shade of other plants (or in some cases rocks) – pioneer species facilitate their existence by providing shade. (The argument has also been made that this type of interaction is exemplary of the tolerance model; see below).
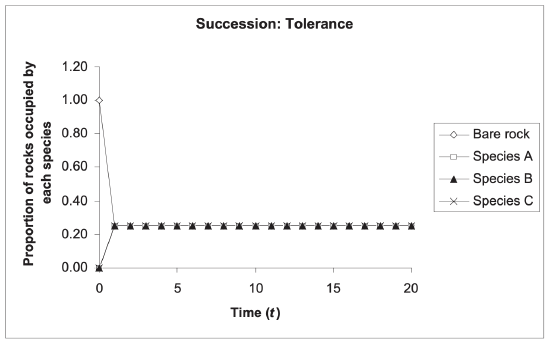
Tolerance model
In the tolerance model , new pioneer species neither inhibit nor facilitate the growth and success of other species. The sequences of succession are thus entirely dependent on life-history characteristics such as the specific amount of energy a species allocates to growth.
- The climax community is composed of the most “tolerant” species that can co-exist with other species in a more densely populated area. Eventually, dominant species replace or reduce pioneer species abundance through competition.
The tolerance model is completely dependent upon life history characteristics. Each species has an equally likely chance to establish itself in the early stages of succession and their establishment results in no environmental changes or impacts on other species (Fig 17.6). Eventually, early species, typically dominated by r-selected species, which prioritize fast rates of reproduction, are out-competed by K-selected species (species that become more dominant when there is competition for limited resources).
For example, we can examine succession in the Loess Plateau in China, where there is initial dominance of the Artemisia scoparia , the pioneer species. Over time, however, the Bothriochloa ischaemum becomes the dominant species and the abundance of A. scoparia greatly declines. This is due to the rapid rate of reproduction of the A. scoparia , resulting in the species’ early abundance, and the dominant competition from the K-selected B. ischaemum , resulting in that species’ later abundance.
A characteristic that is often associated with the tolerance model and well documented in forest succession is survival in conditions of shade. As an uninhabited area becomes populated by different plant species, shade increases – which makes less light available for the next generation. Species that are better adapted to shady conditions will then become dominant.
Reading Question #4
In the tolerance model …
A. Earlier species make the environment more hospitable for other species B. Earlier species make the environment more difficult for other species C. Early species completely prevent other species from establishing D. Earlier species do not directly affect the success of later species
Inhibition model
In the inhibition model earlier successional species inhibit growth of later successional species and reduce growth of colonizing species already present.
- Pioneer species might modify the environment through rapid growth and make the area increasingly shady (essentially increasing competition for light).
- The environment is thus less hospitable to other potential colonizing species.
- The only possibility for new growth/colonization in this successional sequence arises when a disturbance leads to dominating species being destroyed, damaged, or removed. This frees up resources and allows for the invasion of other species that were not previously present
In this model, one species inhibits the presence of another, either through direct means, such as predation (by eating the other species or attacking them), or indirect means, such as competition for resources.
Sometimes in inhibition models, the time of establishment of a species determines which species becomes dominant (Fig 17.7). This phenomenon is referred to as the priority effect and suggests that the species that became established earlier are more likely to become the dominant species. One example of the inhibition model, and the priority effect, occurs in South Australia. In areas where bryozoans are established first, tunicates and sponges cannot grow.
The inhibition model has also been observed at work in forest ecosystems; in these systems the early arrivers hold a monopoly on the land, keeping other species out. Closed shrub canopies have been known to prevent tree growth and access to land for periods of up to 45 years – in an experimental study on inhibition it was found that areas occupied by large areas of Lantana sprawling shrubs excluded and inhibited the growth of tree species.
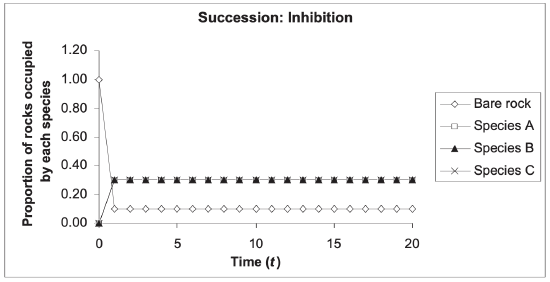
Reading Question #5
In the inhibition model, which species is most likely to become dominant?
A. The species that arrives first B. The species that is the strongest competitor C. The species that is the most shade-tolerant D. The species that can survive with the fewest resources
Adapted from
Clark, M.A., Douglas, M., and Choi, J. (2018). Biology 2e. OpenStax. Retrieved from https://openstax.org/books/biology-2e/pages/1-introduction
Holmberg, T.J. (2022) BIOL 1213. LibreTexts. Retrieved from https://bio.libretexts.org/Sandboxes/tholmberg_at_nwcc.edu/BIOL_1213
change in the environment caused by natural disasters or human activities
also known as early successional species, these are the first species to appear in primary and secondary succession
also known as pioneer species, these are the first species to appear in primary and secondary succession
a hypothesis proposing that maximal species diversity occurs when when environmental disturbance are at an intermediate scale (moderate frequency)
the process by which species in a given area change over time
also known as probiosis, the facilitation model proposes that one species can benefit another species' fitness, and neither species is harmed
this model, in contrast to the facilitation model, proposes that pioneer species do not change abiotic or environmental conditions in a way that affects the invasion of other species
in contrast to the facilitation and tolerance model, this model proposes that early species make it harder for later successional species to inhabit the site and also decrease the growth of other existing species
Introductory Biology 2 Copyright © 2023 by Lisa Limeri is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License , except where otherwise noted.
Share This Book
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Published: 16 October 2012
Regional boreal biodiversity peaks at intermediate human disturbance
- S.J. Mayor 1 ,
- J.F. Cahill Jr 1 ,
- P. Sólymos 1 &
- S. Boutin 1
Nature Communications volume 3 , Article number: 1142 ( 2012 ) Cite this article
4798 Accesses
45 Citations
9 Altmetric
Metrics details
- Biodiversity
- Plant ecology
The worldwide biodiversity crisis has intensified the need to better understand how biodiversity and human disturbance are related. The 'intermediate disturbance hypothesis' suggests that disturbance regimes generate predictable non-linear patterns in species richness. Evidence often contradicts intermediate disturbance hypothesis at small scales, and is generally lacking at large regional scales. Here, we present the largest extent study of human impacts on boreal plant biodiversity to date. Disturbance extent ranged from 0 to 100% disturbed in vascular plant communities, varying from intact forest to agricultural fields, forestry cut blocks and oil sands. We show for the first time that across a broad region species richness peaked in communities with intermediate anthropogenic disturbance, as predicted by intermediate disturbance hypothesis, even when accounting for many environmental covariates. Intermediate disturbance hypothesis was consistently supported across trees, shrubs, forbs and grasses, with temporary and perpetual disturbances. However, only native species fit this pattern; exotic species richness increased linearly with disturbance.
You have full access to this article via your institution.
Similar content being viewed by others

Anthropogenic climate and land-use change drive short- and long-term biodiversity shifts across taxa

Human disturbance is the major driver of vegetation changes in the Caatinga dry forest region

Identifying thresholds in the impacts of an invasive groundcover on native vegetation
Introduction.
The ongoing human-caused global biodiversity decline and continuing expansion of human land use have deepened the need to better understand the relationship of biodiversity to anthropogenic disturbance 1 . The 'intermediate disturbance hypothesis' (IDH), a foundational idea in community ecology, suggests that differences in disturbance among local communities can explain broad patterns in diversity throughout a region 2 , 3 . In communities with moderate disturbance levels, species richness should be higher than in those with low or high disturbance 2 , 3 . After nearly 40 years of research, most studies have found little or no empirical evidence to support the IDH 4 . Support has been even slimmer with anthropogenic than with natural disturbances 4 , 5 , and our ability to predict changes in community diversity patterns following human disturbance is still extremely limited 4 , 5 . Further, studies of the IDH have rarely differentiated native and exotic species, a puzzling oversight given concerns of exotic species invasions, particularly in disturbed environments 6 .
One reason implicated for the scarcity of empirical support for IDH 4 , 5 is that most investigations of diversity–disturbance relationships have been conducted at insufficiently small spatial scales 7 , 8 . Species richness varies non-linearly with spatial scale 9 , 10 , 11 and processes thought to generate IDH vary with scale 12 , so support for the IDH may itself depend on the scale at which it is tested 4 , 7 , 8 . Testing the IDH might best be accomplished by matching the scales of investigation to the scales at which those processes generating the IDH operate. Our understanding of the IDH is poorest at regional extents 4 , 5 (that is, across multiple landscapes), where it has been mostly limited to the system to which IDH was first applied, tropical forest trees. Further, at regional extents—those at which land use planning and management are often conducted—the IDH has generally been tested only with natural not anthropogenic disturbances 4 , 5 .
We tested the specific prediction of the IDH that a non-linear unimodal relationship exists between species richness and anthropogenic disturbance extent, with the greatest number of species present at intermediate disturbance. We conducted our test by examining local communities across the largest spatial extent to date: the 381,047-km 2 boreal ecoregion of Alberta, Canada, an area comparable to Germany. Within this region, vascular plant species richness was sampled with 1 ha surveys of 242 local communities ( Fig. 1 ). We used a direct measure of anthropogenic disturbance: area of land altered by human land use (hereafter 'extent') as assessed with aerial and satellite imagery of 150 m radius circular areas. The sample plots exhibited a complete gradient (0–100%) in human disturbance extent: disturbances varied vastly in intensity, area and type among communities ranging from intact wilderness of one of the world's largest national parks (Wood Buffalo N.P.) to human-dominated landscapes of towns, forestry, agriculture, linear features, conventional oil and gas, and oil sands extraction. We found that species richness of boreal vascular plants showed a unimodal relationship with human disturbance. Richness peaked at intermediate disturbance, with or without accounting for numerous environmental factors, supporting the IDH. Also consistent with the IDH, communities at low and high human disturbance were composed of different sets of species, whereas communities with intermediate disturbance overlapped with those of low and high disturbance. However, we also tested the consistency of the IDH among native and exotic species individually and show that although native species exhibited the predicted peaked relationship of richness to human disturbance, exotic species instead increased linearly.

Presence of each vascular plant species was sampled at 242 sites within the boreal ecoregion of Alberta, Canada. The inset map shows Canada with the boreal region shaded.
Richness–disturbance relationship
A simple unimodal model with peak at 47.7% disturbance best explained vascular plant species richness in relation to human disturbance extent: species richness peaked at intermediate disturbance ( Fig. 2 ), supporting the IDH. That simple quadratic curve ( r 2 =0.135, P <0.001, P of increase in r 2 over linear model <0.001) fit significantly better than other model shapes, whereas the linear regression model was not statistically significant ( r 2 =0.007, P =0.202; Supplementary Table S1 ).

Species richness per hectare of boreal vascular plants as a function of percent total anthropogenic disturbance extent. The black solid curve represents the adopted model, a quadratic curve ( n =242, P <0.001), which supports the IDH. The blue dashed line represents another candidate model, a linear regression that was rejected.
Species composition
The IDH also predicts that composition of species should differ among communities depending on level of disturbance 2 , 3 . Mechanisms thought to drive the peaked disturbance–richness relationship, such as trade-offs in species traits associated with disturbance, suggest that communities with intermediate disturbance are more species rich because they are composed of species associated with both low and high disturbance levels 2 , 3 . Consistent with the IDH, we observed that communities with low (<33.3%) and high (>66.7%) disturbance were composed of significantly different sets of species ( P <0.001), with species composition overlapping least among communities of low and high disturbance ( A =0.028, Fig. 3 ). Also consistent with the IDH, species composition of communities with intermediate (33.4–66.6%) disturbance overlapped with composition of communities with low ( A =0.027) or high ( A =0.017) disturbance ( Fig. 3 ). The difference in composition between intermediate and low disturbance communities was statistically significant ( P <0.001), whereas the difference between intermediate and high disturbance communities was less clear: it was not significantly more different in species composition than would be expected by chance ( P =0.085), but the higher P -value may be due in part to fewer sites in that comparison. Although a high proportion of sites sampled exhibited low disturbance, 30.7% of species were found only at sites with <33.3% human disturbance.

Non-metric multidimensional scaling (NMDS) ordination of species composition across sites. Each point represents a sample site and each site is classified by percent human disturbance (blue: <33.3%, yellow: 33.4–66.6%, red: >66.7%). Overlap of sites in different disturbance levels indicates similarity in species composition between those levels. The axes are unitless.
Species origin and growth form
One possible difference in species composition along the disturbance gradient is the proportion of native and exotic species. These groups are often managed differently: exotic species are traditionally considered of lesser conservation priority than native species and detrimental exotics ('invasive species') are often targets for control 13 . We therefore tested the IDH with native and exotic species separately. We found that although native species richness peaked at intermediate extent of human disturbance ( r 2 =0.104, P <0.001), exotic species richness increased linearly ( r 2 =0.371, P <0.001, P of increase in r 2 for quadratic model over linear=0.082, Fig. 4a ; Supplementary Table S2 ). As native and exotic species showed different shaped functions, we compared the changes in native and exotic richness at ranges in disturbance extent less than or greater than 'intermediate disturbance', where native richness peaked, to better understand relative changes in these species groups. At <47.7% disturbance extent, native species increased (linear regression slope=0.503) more steeply ( P =0.001, ANCOVA) with extent than exotic species (slope=0.119). However, at >47.7% disturbance extent, native species decreased (slope=−0.693, s.e.=0.172) significantly more steeply than exotic species increased (slope=0.022, s.e.=0.038) with disturbance, as indicated by the standard errors of the absolute values of the slopes that were less than the gap between those errors. Anthropogenic disturbance was a more accurate predictor of richness of exotic species than of natives; disturbance extent explained 3.6 times the variance in exotics than in natives ( Fig. 4a ).

Comparison of species richness per hectare of native and exotic boreal vascular plants in relation to percent anthropogenic disturbance extent. ( a ) Native (black circles) and exotic (blue squares) species richness as a function of disturbance extent. A quadratic model fit (solid black curve) native species richness with its peak at intermediate disturbance ( n =237, P <0.001), whereas a linearly increasing model (dashed blue line) fit exotic species richness over human disturbance ( n =237, P <0.001). ( b ) Ratio of native to exotic species richness as a function of disturbance extent decreased linearly.
The ratio of native to exotic species declined linearly with human-disturbed area (slope=−0.247, r 2 =0.203, P <0.001, Fig. 4b ). The IDH was conceived without apparent consideration of invasive species, and the non-conformance of exotics implies the hypothesis may only apply to regional metacommunities with a 'closed' species pool. The correlation of exotic species and human disturbance is consistent with previous studies 13 , and suggests that fundamentally different responses to disturbance exist among some groups. However, when we categorized species by 'growth form' 14 , another common ecological categorization and indicator of functional role, richness of each type of vascular plant species (including forbs, graminoids, shrubs and trees), peaked at intermediate disturbance extent ( Supplementary Table S3 ). The generality of the IDH among types of species requires greater scrutiny.
Environmental covariates
Total species richness varied considerably along the disturbance gradient, an expected result shared by other related observational studies 4 , 5 , 15 owing to effects of numerous other factors including climate, topography, stand age, soil and other environmental conditions. We accounted for many such factors by constructing multiple regression models that included 19 environmental variables ( Supplementary Table S4 ) including human disturbance, and again compared possible shapes of the relationship between richness and human disturbance. Consistent with the IDH, a quadratic curve with maximum species richness at intermediate disturbance fits best ( P <0.001, Supplementary Table S5 ). As expected, including the environmental variables in the model better explained species richness across sites ( r 2 =0.709) than did the simple quadratic model of human disturbance in response to richness ( r 2 =0.137, P =0.780; Supplementary Table S5 ). However, accounting for these variables did not qualitatively change the shape of the relationship between richness and disturbance.
Anthropogenic disturbance types
Anthropogenic disturbance varied greatly in intensity, frequency, permanence and spatial characteristics, so different types of disturbance might be expected to drive different disturbance–diversity relationships as a result. Agricultural disturbance is often so frequent and intensive (for example, annual cultivation) such that successional processes are minimal, and primary roads plus urban and industrial sites are virtually permanent landscape changes. We classified those disturbances as 'perpetual'. By contrast, vegetation on forestry cut blocks, pipelines and seismic lines usually exhibit dynamic and successional changes in species composition following initial disturbance, classified here as 'temporary'. To investigate the relationship of richness to proportion of temporary human footprint, we excluded perpetual human footprint. In areas not perpetually disturbed, richness again peaked at intermediate temporary disturbance extent. The simple quadratic model with peak at 55.3% temporary disturbance fit significantly better than other model shapes ( r 2 =0.1136, P <0.001, P of increase in r 2 over linear model <0.001; Supplementary Table S6 ); the linear model had a likelihood of 0.011 (ΔAIC=9.05, AIC weight=0.008). Thus, the IDH was supported both for total disturbance (including perpetual disturbance) and for temporary disturbance alone, suggesting the hypothesis is robust to disturbance permanence.
Our results for native species in anthropogenically disturbed communities lend support to the IDH, in contrast to more than 80% of other studies that did not exhibit the predicted unimodal relationship 4 , 5 . We attribute support for IDH found in this study but not many others to several aspects of study design. First, our study was conducted at a large regional extent. Reviews of IDH concluded many tests of IDH have been inadequately small scaled 4 , 5 , 8 ; few tests have approached the regional extents at which the IDH was intended to apply (for example, the 35,000-ha Budongo rainforest 3 ). Connell 3 , who advanced the IDH, was specific in stating this hypothesis applied to 'variation in diversity among local stands (that is, communities)', not among sites within small communities nor among large biomes. Diversity–disturbance relationships at either local (for example, within a single community) or interregional scales are generated by different processes and tend to exhibit different diversity patterns 16 . Our study was truly regional because we observed consistent results among widely separated communities sampled across a large geographic range. Second, we utilized a direct measure of disturbance: area altered by human land use. Data-rich studies of tropical forest trees 8 , 17 , 18 have used indirect proxies for disturbance such as % pioneer species or canopy height, which may have contributed to their opposing conclusions and which have been criticized for their poor interpretability 7 . Third, the communities under investigation varied on a full gradient of 0–100% disturbance extent. Many studies may have encompassed a narrow range of disturbance levels, thereby risking missing any 'intermediate' peak in richness 4 , 5 , 8 .
Our results have important implications for prominent theories of species invasion, which predict a strong link between richness of native and exotic species. Communities richer in native species may be more resistant to invasion of exotics due to competition among species, thereby producing a negative relationship between native and exotic richness 19 , 20 , 21 . A contrasting idea suggests native and exotic species richness should be positively related because the key environmental factors driving richness are shared by native and exotic species 22 , 23 . We found little support for either of these conflicting predictions: native species ( x ) weakly explained exotic ( y ) richness ( y =0.030 x +1.440, r 2 =0.033, P =0.005). By contrast, human disturbance was a relatively good predictor of exotic richness ( r 2 =0.371, P <0.001), supporting the view that exotic species are not independent drivers of native biodiversity declines, but 'passengers' simply accompanying community change in disturbed environments 24 .
Considerations of biodiversity conservation in regional land use planning decisions usually focus on preservation of species, which are at risk or have particular economic or cultural value. However, biodiversity is increasingly being more broadly considered in land use planning decisions 25 , species richness is considered in conservation prioritization 26 and ecological theory is being called upon to aid conservation-oriented land use decisions 27 . We show that the IDH can partially explain the variation in species richness found throughout a large region subjected to a diversity of anthropogenic impacts. Although simple, IDH is an enduring idea that may be applicable to land use planning 28 and along with invasion theories can contribute to understanding changes in native richness expected with human land use 29 . Our results are generally consistent with both the IDH and the more specific 'patch mosaic dynamics hypothesis', which suggests that landscapes with a mosaic of disturbed patches of various intensity, extent and frequency can host the greatest diversity 30 . However, our findings also suggest that management strategies designed to maximize biodiversity by imposing intermediate human disturbance regimes could result in communities with species composition statistically indistinguishable from those at high disturbance ( Fig. 3 ), exclusion of some species or community types found only at low disturbance, and proportionally fewer native species 29 ( Fig. 4b ). Thus, we strongly suggest any management application of this work takes into consideration the overall species composition across the study area, and not focus solely on species richness levels in individual plots. As disturbance was more strongly correlated with and better predicted richness of exotics than natives, management efforts aimed exclusively at maximizing species richness could have a greater impact on exotic species than natives. We found that although human disturbance significantly predicted native species richness ( r 2 =0.104, P <0.001), the interactive effects of disturbance and exotic species richness explained nearly twice as much variation in native richness as did disturbance alone ( r 2 =0.206, P <0.001). Efforts to conserve native species diversity should therefore consider not only management of human disturbance but also the interactions of land use and exotic species, such as the roles of human disturbance in creating opportunities for exotic species establishment or acting as conduits for exotic dispersal 29 , 31 . Catford et al . 29 suggest a range of management options informed by IDH and invasion patterns. However, the inconsistency in support and possible scale dependence of IDH warrant caution in its application. In the boreal biome, among the world's largest and most intact terrestrial ecoregions, the IDH may serve as a starting point for understanding how continued expansion of human activities may affect local plant communities.
Study design
Vascular plant richness was surveyed in the boreal ecoregion of the province of Alberta, Canada by the Alberta Biodiversity Monitoring Institute, using standardized protocols 32 . Vascular plant species occupancy was surveyed within 1 ha for 25 min at 242 sites, a subset of 959 sites of the Canadian National Forestry Inventory grid with grid centre spacing of 20 km ( Fig. 1 ). All surveys were conducted from 26 June to 18 August of 2003–2009. Human disturbance extent (% land area converted by humans) was assessed by manual interpretation of 1:30,000 aerial photos and SPOT satellite imagery within circles of 150 m radius (7.07 ha) at each site. Disturbance extent was assessed within a larger area than that at which richness was surveyed, because both direct local disturbances and those in the surrounding landscape may potentially affect local richness 33 . We distinguished native and exotic species according to ACIMS 34 .
To determine the best-fit shape of the richness–disturbance relationship, we performed linear and polynomial regression analyses. Polynomial (quadratic and cubic) models required additional model parameters not included in simple linear regression, so we adopted those models only when they were both significant (reported as ' P <0.05') and when they fit significantly better by explaining significantly more variation than the simpler model of fewer parameters, as diagnosed with an ANOVA of candidate models (reported as ' P of increase in r 2 over linear model'). In all cases, that frequentist model selection protocol resulted in the same adopted model as with model selection by AIC (highest model likelihood, AIC weight >0.5).
Disturbance types
Human disturbance only described spatial extent of disturbance; it was not a measure of 'area removed' (as in some theoretical models) and we did not quantify intensity, frequency or time as disturbance, which varied greatly within and among disturbance types. Anthropogenic disturbances included agricultural pasture and croplands, forestry cut areas of variable ages, linear features (including pipelines, powerlines, roads, railways and cutlines primarily for oil/gas seismic exploration), industrial/commercial disturbances (including coal and mineral surface mines, oil and gas well pads, communication towers, gravel pits, heavy oil sands development and spoil piles), and both urban and rural settlements. Natural disturbances such as fire and insect damage were not correlated to anthropogenic disturbance.
To determine if species composition of communities was consistent with proposed mechanisms for the IDH, we tested for differences in species composition among communities varying in human footprint. We classified sites as <33.3, 33.4–66.6 and >66.7 human footprint and used a multi-response permutation procedure using a Euclidean distance measure and rank-transformed distance matrix to make pairwise comparisons of species composition across sites grouped by human footprint class. We performed non-metric multidimensional scaling analysis, an ordination technique that graphically presents the overlap in composition across sites, then classified each site by disturbance level ( Fig. 3 ).
We used a common non-taxonomic categorization of plant species—'growth form'—to explore the consistency of the IDH among species of different functional types 14 . Vascular plant species in boreal Alberta vary from small herbaceous forbs to tall woody canopy trees, and because these different growth forms have different functional roles, they could be expected to respond differently to disturbance 35 .
To determine the best-fit shape of the richness–disturbance relationship while accounting for potentially confounding variables, we constructed multiple regression models and selected model shapes as above. For human disturbance and each environmental covariate, we selected the best fitting linear or polynomial form. We used a subset of 146 sites for which data was available for 18 potentially confounding variables. We included in the models human disturbance plus the following: natural subregion type, latitude, longitude, elevation, topographic heterogeneity, growing degree days, mean annual temperature, mean annual precipitation, terrain wetness, site wetness, solar flux, canopy closure, oldest tree age, organic depth, soil type, surficial geology, slope position and landform classification ( Supplementary Table S4 ). Too few data were available to include natural disturbance (year of last fire and natural disturbance extent) in the models. Details of environmental variable observation and estimation are available at www.abmi.ca .
Temporary anthropogenic disturbance
To investigate the relationship of richness to temporary disturbance, we categorized forestry cut areas, cutlines, pipelines and powerlines as temporary, and all other disturbances as perpetual. We first excluded all sites with any perpetual disturbance at the 1-ha scale where vascular plants were sampled. We then subtracted any perpetually disturbed area in the 150-m radius circle plot and calculated the proportion of temporary disturbance in the remaining area.
Additional information
How to cite this article: Mayor, S.J. et al . Regional boreal biodiversity peaks at intermediate human disturbance. Nat. Commun. 3:1142 doi: 10.1038/ncomms2145 (2012).
Barnosky, A. D. et al. Has the Earth's sixth mass extinction already arrived? Nature 471 , 51–57 (2011).
Article CAS ADS Google Scholar
Grime, J. P. Competitive exclusion in herbaceous vegetation. Nature 242 , 344–347 (1973).
Article ADS Google Scholar
Connell, J. H. Diversity in tropical rain forests and coral reefs. Science 199 , 1302–1310 (1978).
Mackey, R. L. & Currie, D. J. The diversity-disturbance relationship: is it generally strong and peaked? Ecology 82 , 3479–3492 (2001).
Google Scholar
Shea, K., Roxburgh, S. H. & Rauschert, E. S. J. Moving from pattern to process: coexistence mechanisms under intermediate disturbance regimes. Ecol. Lett. 7 , 491–508 (2004).
Article Google Scholar
Hobbs, R. J. & Huenneke, L. F. Disturbance, diversity, and invasion: implications for conservation. Conserv. Biol. 6 , 324–337 (1992).
Sheil, D. & Burslem, D. F. R. P. Disturbing hypotheses in tropical forests. Trends Ecol. Evol. 18 , 18–26 (2003).
Bongers, F., Poorter, L., Hawthorne, W. D. & Sheil, D. The intermediate disturbance hypothesis applies to tropical forests, but disturbance contributes little to tree diversity. Ecol. Lett. 12 , 798–805 (2009).
Preston, F. W. The canonical distribution of commonness and rarity: Part I. Ecology 43 , 185–215 (1962).
MacArthur, R. H. & Wilson, E. O. In The Theory of Island Biogeography (Princeton Univ Press, Princeton, NJ, 1967).
Whittaker, R. J., Willis, K. J. & Field, R. Scale and species richness: towards a general, hierarchical theory of species diversity. J. Biogeogr. 28 , 453–470 (2001).
Cadotte, M. W. Competition-colonization trade-offs and disturbance effects at multiple scales. Ecology 88 , 823–829 (2007).
Davis, M. A. In Invasion Biology (Oxford University Press, USA, 2009).
USDA-NRCS. The PLANTS Database (2011).
Hughes, R. A., Byrnes, J. E., Kimbro, D. L. & Stachowicz, J. J. Reciprocal relationships and potential feedbacks between biodiversity and disturbance. Ecol. Lett. 10 , 849–864 (2007).
Sax, D. F. & Gaines, S. D. Species diversity: from global decreases to local increases. Trends Ecol. Evol. 18 , 561–566 (2003).
Hubbell, S. P. et al. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 283 , 554 (1999).
Molino, J. F. & Sabatier, D. Tree diversity in tropical rain forests: a validation of the intermediate disturbance hypothesis. Science 294 , 1702 (2001).
Elton, C. S. In The Ecology of Invasions by Animals and Plants (Methuen & Co. Ltd, London, UK, 1958).
Kennedy, T. A. et al. Biodiversity as a barrier to ecological invasion. Nature 417 , 636–638 (2002).
Tilman, D. Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proc. Natl Acad. Sci. USA 101 , 10854 (2004).
Stohlgren, T. J., Barnett, D. T. & Kartesz, J. T. The rich get richer: patterns of plant invasions in the United States. Front. Ecol. Environ. 1 , 11–14 (2003).
Levine, J. M., Adler, P. B. & Yelenik, S. G. A meta analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 7 , 975–989 (2004).
MacDougall, A. S. & Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 86 , 42–55 (2005).
Boutin, S., Haughland, D. L., Schieck, J., Herbers, J. & Bayne, E. A new approach to forest biodiveristy monitoring in Canada. For. Ecol. Manage. 14 , S168 (2009).
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403 , 853–858 (2000).
Bestelmeyer, B. T., Miller, J. R. & Wiens, J. A. Applying species diversity theory to land management. Ecol. Appl. 13 , 1750–1761 (2003).
Sasaki, T. et al. Management applicability of the intermediate disturbance hypothesis across Mongolian rangeland ecosystems. Ecol. Appl. 19 , 423–432 (2009).
Catford, J. A. et al. The intermediate disturbance hypothesis and plant invasions: Implications for species richness and management. Perspect. Plant Ecol. 5 , 231–241 (2012).
Pickett, S. T. A. In The Ecology of Natural Disturbance and Patch Dynamics (Academic Pr, 1986).
Didham, R. K., Tylianakis, J. M., Gemmell, N. J., Rand, T. A. & Ewers, R. M. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol. Evol. 22 , 489–496 (2007).
ABMI. Alberta Biodiversity Monitoring Institute, Terrestrial field data collection protocols (Edmonton, Canada, 2009). Available at: ( http://www.abmi.ca ).
Turner, M. Disturbance and landscape dynamics in a changing world. Ecology 91 , 2833 (2010).
ACIMS. Alberta Conservation Information Management System . Alberta Tourism Parks and Recreation. (Edmonton, Canada, 2010).
Lavorel, S., McIntyre, S., Landsberg, J. & Forbes, T. Plant functional classifications: from general groups to specific groups based on response to disturbance. Trends Ecol. Evol. 12 , 474–478 (1997).
Article CAS Google Scholar
Download references
Acknowledgements
Raw data was provided by Alberta Biodiversity Monitoring Institute following their established data collection protocols and is available at www.abmi.ca . Funding was provided by Natural Sciences and Engineering Research Council of Canada (Discovery Grants to J.F.C., S.B.; Canada Graduate Scholarship to S.J.M.), Alberta Ingenuity Fund (S.J.M.), Alberta Biodiversity Monitoring Institute (S.J.M.), Alberta Sport Recreation Parks & Wildlife Foundation (S.J.M.) and Canadian Circumpolar Institute (S.J.M.).
Author information
Authors and affiliations.
Department of Biological Science, University of Alberta, Edmonton, T6G 2E9, Alberta, Canada
S.J. Mayor, J.F. Cahill Jr, P. Sólymos & S. Boutin
Department of Renewable Resources, University of Alberta, Edmonton, T6G 2H1, Alberta, Canada
You can also search for this author in PubMed Google Scholar
Contributions
S.J.M., J.F.C., S.B. and F.H. conceived, designed and supervised the study, and edited multiple versions of the manuscript. S.J.M. performed data analyses and led the writing. S.J.M. and P.S. designed and executed the temporary disturbance analyses.
Corresponding author
Correspondence to S.J. Mayor .
Ethics declarations
Competing interests.
The authors declare no competing financial interests.
Supplementary information
Supplementary information.
Supplementary Tables S1-S6. (PDF 63 kb)
Rights and permissions
Reprints and permissions
About this article
Cite this article.
Mayor, S., Cahill, J., He, F. et al. Regional boreal biodiversity peaks at intermediate human disturbance. Nat Commun 3 , 1142 (2012). https://doi.org/10.1038/ncomms2145
Download citation
Received : 25 June 2012
Accepted : 20 September 2012
Published : 16 October 2012
DOI : https://doi.org/10.1038/ncomms2145
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
Intermediate disturbance hypothesis explains eutrophication and biodiversity pattern in a boreal river basin, china.
- Shuang Tian
Hydrobiologia (2022)
Singing in the suburbs: point count surveys efficiently reveal habitat associations for nocturnal Orthoptera across an urban-to-rural gradient
- Darin J. McNeil
- Christina M. Grozinger
Journal of Insect Conservation (2020)
Structure and composition of tree species in tropical moist deciduous forests of Eastern Ghats of Odisha, India, in response to human-induced disturbances
- Tirthabrata Sahoo
- Laxmikanta Acharya
- Pratap Chandra Panda
Environmental Sustainability (2020)
Long-term grazing impacts on vegetation diversity, composition, and exotic species presence across an aridity gradient in northern temperate grasslands
- Mark P. Lyseng
- Edward W. Bork
- Walter D. Willms
Plant Ecology (2018)
Global terrestrial Human Footprint maps for 1993 and 2009
- Oscar Venter
- Eric W. Sanderson
- James E.M. Watson
Scientific Data (2016)
By submitting a comment you agree to abide by our Terms and Community Guidelines . If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

We can help you reset your password using the email address linked to your BioOne Complete account.

- BioOne Complete
- BioOne eBook Titles
- By Publisher
- About BioOne Digital Library
- How to Subscribe & Access
- Library Resources
- Publisher Resources
- Instructor Resources
- FIGURES & TABLES
- DOWNLOAD PAPER SAVE TO MY LIBRARY
The intermediate disturbance hypothesis (IDH) suggests that the peak of species diversity occurs at intermediate-scale disturbances. The IDH received criticisms because many studies have shown that the relationship between disturbances and species diversity is generally not unimodal. We searched Web of Science for articles on IDH to study the applications of the hypothesis in animal and plant studies. We classified found articles into those which presented evidence in favour and against the IDH. Furthermore, we analysed the effects of article age and impact factor of the journal in which it was published on the number of citation this article received. We found that most arguments against the IDH were found in papers on aquatic ecology and in papers published in journals with higher impact factors. Those articles were also cited more often than those presenting evidence in support of the IDH. We thus can conclude that the IDH seems to be less supported in newer papers and particularly in those in the field of aquatic ecology.

KEYWORDS/PHRASES
Publication title:, collection:, publication years.
The Intermediate Disturbance Hypothesis and the Diversity of Bacteria on the Human Body

Patrick Hanes Department of Biology Lake Forest College Lake Forest, Illinois 60045 Download PDF
The intermediate disturbance hypothesis predicts that the highest diversity will occur at levels of moderate disturbance. We measured the species diversity, abundance, and species diversity of bacteria on the human body in relation to disturbance by washing. Bacteria were sampled from three surfaces on the human body with varying disturbances, counted, and classified into morphospecies. The site with intermediate disturbance had the highest species richness and species diversity. While these results match the prediction of the intermediate disturbance hypothesis, diversity of bacteria may have also been influenced by physical and behavioral factors.
Introduction
The intermediate disturbance hypothesis predicts that the greatest diversity occurs at moderate levels of disturbance. High levels of disturbance will wipe out all species, and low levels of disturbance allow superior competitors to eventually drive other species to extinction (Jiang and Patel, 2008; Kadmon and Bengamini, 2006; Roxburgh et al., 2004; Shea et al., 2004). Disturbances can include alterations to the environmental structure, habitat fragmentation, and biotic factors, and can simply be defined as anything that changes the composition of niches (Shea et al., 2004). These disturbances increase the availability of resources to new species by eliminating or reducing the populations of potential competitors (Jiang & Patel, 2008).
The species richness of a post-disturbance environment is due to a combination of factors. Disturbance only promotes greater species richness if it occurs in between the generation time of the slowest-reproducing species and the time needed for complete dominance by the best competitor (Shea et al., 2004). The area of a disturbed environment brings in the factor of dispersal versus competition (Roxburgh et al., 2004; Shea et al., 2004). Both the intensity and duration of the disturbance will have different effects on each species and life history stage, which may range from no effect to the death of an individual (Shea et al., 2004). Productivity relative to the severity of disturbance could affect the species richness of the post-disturbance environment by influencing the population sizes and competitive advantages for native species (Kadmon & Benjamin, 2006).
Disturbed environments undergo a transition as recovery proceeds (Roxburgh et al., 2004). The intermediate disturbance hypothesis assumes that diversity is driven by species adapting to the trade-off between tolerating fluctuations in environmental conditions and competing for resources (Jiang & Patel, 2008; Kadmon & Benjamini, 2006). Due to these differences in adaptation, the potential coexistence would not be completely determined by competitive advantages and may come about by a variety of mechanisms (Roxburgh et al., 2004). Resulting species composition and richness were also shown to be influenced by the order in which new species colonized the post-disturbance environment (Jiang & Patel, 2008).
Bacteria on the human body are subjected to many of the same factors that play into the species richness under the intermediate disturbance hypothesis, such as exposure to disturbances of varying intensity and duration, colonization and degrees of tolerance to different environmental stresses (Fierer et al., 2008; Grice & Segre, 2011). While bacterial communities have been shown to differ between sites on the human body, the factors that determine this diversity are still poorly understood (Fierer et al., 2008).
In order to test the intermediate disturbance hypothesis, we studied the bacteria living on similar exposed surfaces of the human body of male and females under the disturbance of washing. The maximum disturbance was represented by the back of the hands, which are subjected to constant scrubbing and exposure to soap. Minimal disturbance was represented by the inner forearms, which are usually disturbed when washed once a day while showering. It was thought that the cheek lies somewhere in between these two extremes, usually being subjected to a moderate amount of disturbance of washing just a few times a day. Therefore, cheeks should show significantly higher species richness than the back of the hands or forearms. All three of these surfaces are characterized as dry, external environments. We also predicted there will be no differences in diversity between males and females of these surfaces. We sampled these areas of the body to determine patterns of species richness, abundance, and species diversity on the human body.
We sampled 20 young adults, 10 males and 10 females, at Lake Forest College in late March and early April of 2013. Q-tips were rubbed thoroughly on areas of interest (back of the hand, inner forearm, and cheek) for approximately 5 seconds in order to collect bacteria. It was noted when the participants last showered or if they wore make up or had facial hair on the sampled surface, as these factors may affect the growth and community composition of bacteria. Samples were collected from the same side of the body. We swirled the contaminated end of the Q-tip in a microfuge tube filled with 0.5 mL of PBS solution and chilled the tubes in a refrigerator for 24 hours. Each tube was then shaken and 50 μL of solution were pipetted out and diluted into a second microfuge tube filled with 0.95 mL of solution. We pipetted 50 μL of solution from the second tube and applied it to the surface of a TSA plate. The mixture was evenly spread over the plate surface with 5-7 sterile glass beads and left to incubate at 36°C for 6 days.
We counted and classified the grown bacteria into morphospecies by the color and texture of the colonies. We used a 2-way Anova to test for differences between areas for species richness, abundance, and species diversity. Species diversity was calculated by using the Shannon-Weiner Diversity Index.
We observed 621 colonies comprised of 15 morphospecies (Fig. 1). Two morphospecies classified as “white, shiny” and “grey, dull” were the most abundant, comprising 17.7% and 60.3% of colonies respectively. Only one species was represented by just one colony. 14 of the 60 plates exhibited no growth of bacterial colonies. Of our participants, seven of the females were wearing makeup, four men had facial hair, and 13 showered the morning of sampling, while five showered the morning before.
The average for the species richness was what would be expected if they were driven by the intermediate disturbance hypothesis. The check had the highest average number of species per sample (2.75±0.29 species) compared to the back on the hand and the forearm (Fig. 2). Males with facial hair had higher mean species richness on the cheek (3.75±1.43). Females without make up had higher mean species richness (6±1.21). Participants who showered the morning before being sampled had higher mean species richness (2±0.75) compared to those who showered the morning of sampling (1.95±0.55). Species richness exhibited a significant difference for the model (F5, 60=2.55, P=0.049), sex (P=0.019), and surface (P=0.045). There was no significance for the interaction of the sex and surface (P=0.875).
The mean abundance of bacterial colonies was greatest on the female hand (30.4±26.05) (Fig. 3). However, this data was skewed by an outlier. When the outlier was removed, the average abundance of colonies on the hand was reduced to 1.78 per sample, and the male cheek had the highest average abundance (12.5±5.23). Males with facial hair had higher mean abundance on the cheek (21.25±12.45). Females wearing makeup had a slightly higher mean abundance on the cheek (3.71±1.69). Participants who showered the morning of sampling had greater mean abundance of colonies (12.90±12.70) than those who showered the morning before sampling (6.36±6.74). Abundance was insignificant for the model (F5, 60=0.720, P=0.611), sex (P=0.550), surface (P=0.586), and the interaction of the sex and surface (P=0.347).
Species diversity was greatest on the cheeks of both males and females. The H values of the Shannon-Weiner Index for males and females were (2.41±0.04) and (1.91±0.03) respectively (Fig. 4). Species diversity was significant for the model (F5, 90=2.551, P=0.34), sex (P=0.049), and surface (P=0.017). There was no significance for interaction of the sex and surface (P=0.898).

Figure 1 : A sample of cultures. Clockwise from the top left: hand, forearm, cheek, and no culture present.

Figure 2: The mean species richness per sample for surface and sex. The cheek exhibited the greatest species richness for both sexes. Discussion

Figure 3: The mean abundance of colonies per sample for surface and sex. There was no observable trend.

Our predictions were supported for species richness and species diversity, both of which follow the pattern for the intermediate disturbance hypothesis. We fail to reject the null hypothesis for the abundance of colonies for each surface. We also fail to reject the null hypothesis for the abundance differences by sex. The average for all measurements showed initial differences among the three surfaces. While the cheek was highest in average species richness and species diversity, the variation the variation of measurements between the hand and forearm could be due to environmental differences other than the level of disturbance. Differences between sexes in these measurements were apparent, and may be due to physical or behavioral factors. At intermediate disturbance, the gap in measurements between males and females closes, which means disturbance at this level may cancel out any effects that contribute to differences in sex.
The removal of the outlier for abundance made the made the male cheek have the greatest abundance of bacterial colonies. This inconsistency and lack of significance between surfaces for abundance may be due to several factors. Disturbances have a unique effect on each species, of which some will suffer greater reduction in their populations than others (Roxburgh et al., 2004; Shea et al., 2004). Therefore, the insignificant differences in the bacterial colonies could be due to the abilities of a dominant species being compromised by disturbance, leading to a more equal distribution of populations. The opposite trends in the abundance for males and females among surfaces are possibly due any differences in the environment dependent on sex.
Subtle differences in the physical traits of areas of the human body could affect the factors that shape the intermediate disturbance hypothesis. The skin of a human can be categorized into habitats by the presence of hair follicles, secretion of salt and water, temperature, humidity, and topography (Costello et al., 2009; Grice & Segre, 2011). These factors were not accounted for in our study and almost certainly have influence on bacterial communities. The dry surfaces of the human body, including the forearm, tended to be more diverse (Grice & Segre, 2011). The diversity of the forearm was usually higher than the hand, especially for men. Men tend to have more acidic skin and produce more sebum, both of which tend to lower bacterial diversity (Fierer et al., 2008). However, males were higher in our measurements of species diversity, so this doesn’t seem to be a viable cause. Yet, other sex-specific factors, such as the production of sweat and hormones, may be possible factors (Fierer et al., 2011).
The presence of either facial hair or makeup over the cheek may have created a different microhabitat than the arm or hand and could alter the effects of disturbance. Facial hair could act as a barrier to minimize the disturbance of washing and present more of a heterogeneous environment. Stability and structural heterogeneity have been attributed to an increase in species richness by allowing organisms to evolve into new forms with little extinction and specialize within a structurally varied environment (Pianka, 1966). Makeup could act as a barrier that traps heat and moisture to the underlying skin. High temperatures and humidity are said to promote diversity, and environments of the human body with these traits were found to have greater quantities of bacteria (Grice & Segre, 2011; Pianka, 1966).
Behavior of participants could affect the diversity of bacteria at the time of sampling. Time between disturbances is a factor in the ability of a community to persist through a disturbance (Shea et al., 2004). This factor is represented by the how long it had been since the participants showered before sampling. The time since hand washing has a significant effect on the community composition of bacteria, but not diversity, and females were more likely to have washed their hands recently (Fierer et al., 2008). The mean species richness of our participants increased with time since they last showered, while greater abundance was observed from participants who showered the morning of sampling. However, it is unknown what effects hand washing had on the hands of our participants, as they all sampled at least an hour after washing their hands. Frequencies of hand washing have been observed to differ by sex and may explain the differences in diversity between males and females. We assumed the surfaces sampled for bacteria came into negligible instances of contact with foreign surfaces. Any increased contact with foreign surfaces would offer more opportunities for bacteria to colonize a post-disturbance environment and establish themselves in the new location (Fierer et al., 2008; Jiang & Patel, 2008).
Understanding the factors that drive patterns of diversity for bacteria on human skin may hold the potential for the treatment and prevention of skin disease. Alterations to the composition of bacterial communities on the skin have been known to cause infections and disorders (Grice & Segre, 2011). Washing as a means of hygiene has been shown to be a possible mechanism for the propagation of bacterial diversity. This resulting diversity may influence the ability of a pathogen to infect a host. The opening of niche space that arises out of a disturbance facilitates the establishment of new colonizers to a community, and it is possible that this may also apply to pathogens (Jiang & Patel, 2008). However, greater diversity from intermediate disturbance may also suppress species that lead to disease (Grice & Segre, 2011). If the effects of high skin bacteria on pathogen colonization can be determined, it may be possible to use our skin bacteria as a form of biological control that can evolve with diseases. In order to truly further our understanding of skin disease, normal states of bacterial communities must first be ascertained (Costello et al., 2009; Fierer et al., 2008).
A more accurate pattern of bacterial diversity on the human body may be determined in future studies. A larger sample size would compensate for any outliers and could provide a more complete collection of bacterial species that occur on the human body. Other variables, such as facial hair or makeup, sex, and time since showering should be held constant to eliminate their possible effects on the response of bacteria to disturbance. Comparisons between variants of these factors would make a great study for the types of environment that lead to diversity and if diversity is more likely to be the result of stability or disturbance. Understanding these factors may explain why surfaces with the most and least disturbance still showed differences in diversity. Washing seemed to act as an adequate disturbance, but it is worth investigating what other types of disturbances, such as chemical or abrasive, could result in the highest diversity at intermediate levels.
Cardotte, M.W. (2007). Colonization-competition trade-offs and disturbance effects at multiple scales. Ecology, 88, 823-829.
Costello, E. K., Lauber, C. L., Hamady M., Feier, N., Gordon, J. I., & Knight, R. (2009). Bacterial community variation in human body habitats across space and time. Science, 326, 1694-1697.
Feier, N., Hamady, M., Lauber, C. L., & Knight, R. (2008). The influence of sex, handedness, and washing on the diversity of hand surface bacteria. Proceedings of the National Academy of Sciences of the United States of America, 105, 17994-17999.
Grice, E. A., & Segre, J. A. (2011). The skin microbiome. Nature Reviews Microbiology, 9, 244-253. Jiang, L., & Patel S. N. (2008). Community in the presence of disturbance: a microcosm experiment. Ecology, 89, 1931-1940.
Kadmon, R., & Benjamini, Y. (2009). Effects of productivity and disturbance on species richness: A neutral model. The American Naturalist, 167, 939-946.
Pianka, E. R. (1966). Latitudinal gradients in species diversity: A review of concepts. The American Naturalist, 100, 33-45.
Roxburgh, S. H., Shea, K., & Wilson, J. B. (2004). The intermediate disturbance hypothesis: Patch dynamics and mechanisms of coexistence. Ecology, 85, 359-371.
Shea, K., Roxburgh, S.H., & Rauschert, E. S. J. (2004). Moving from pattern to process: coexistence mechanisms under intermediate disturbance regimes. Ecology Letters, 7, 491-508.
objects/intermediate-disturbance-mosaic.html
last edited on November 8, 2011 by John Rueter
COMMENTS
The intermediate disturbance hypothesis ( IDH) suggests that local species diversity is maximized when ecological disturbance is neither too rare nor too frequent. At low levels of disturbance, more competitive organisms will push subordinate species to extinction and dominate the ecosystem. [1] At high levels of disturbance, due to frequent ...
On October 26, 2020October 26, 2020 By outoftheblueocean. The Intermediate Disturbance Hypothesis is one of the staples of ecology, especially marine ecology. The Intermediate Disturbance Hypothesis was first proposed by Connell, a well-known intertidal and general ecologist, in 1978 (See his article "Diversity in Tropical Rain Forests and ...
The Intermediate Disturbance Hypothesis☆ R.W. Osman, in Encyclopedia of Ecology (Second Edition), 2015 Gradients and Landscapes. The intermediate disturbance hypothesis has been applied and tested in larger systems, in particular to spatial patterns of diversity along gradients and within landscapes. Gradients and zonation patterns often have communities dominated by different species at ...
Intermediate Disturbance Hypothesis. Disturbance variability and species diversity are heavily linked. The success of a wide range of species from all taxonomic groups is closely tied to natural disturbance events such as fire, flooding, and windstorm. Diversity tends to be low in harsh environments because of the intolerance of all but ...
The intermediate disturbance hypothesis (IDH) and the dynamic equilibrium model (DEM) are influential theories in ecology. The IDH predicts large species numbers at intermediate levels of disturbance and the DEM predicts that the effect of disturbance depends on the level of productivity. However, various indices of diversity are considered ...
intermediate disturbance hypothesis The proposition that the highest diversity of species in an ecosystem is maintained by a level of disturbance intermediate between frequent and rare disturbance. If disturbance is frequent the succession may fail to develop beyond the pioneer phase.If disturbance is rare, the climax will be established and diversity reduced according to the competitive ...
The realisation that the frequency of disturbance can influence community structure led to the formulation of the intermediate disturbance hypothesis (IDH) (Connell, 1978; Figure 22 below). The IDH proposes that species diversity is generally maximised if disturbance is neither too rare nor too frequent because species that thrive at both early ...
The intermediate disturbance hypothesis (IDH) has been used for several decades as an explanation for the coexistence of species in ecological communities. It is intuitively simple, but deceptively so. We show, via discussion and examples, that the IDH is not one mechanism of coexistence, but rather summarizes a set of similar phenomena that ...
Disturbance Hypothesis. An influential ecological theory, the intermediate disturbance hypothesis (IDH), predicts that intermediate levels of disturbance will maximize species diversity. Empirical studies, however, have described a wide variety of diversity-disturbance relationships (DDRs). Using experimental populations of microbes, we show ...
The intermediate disturbance hypothesis (IDH) has been used for decades as an explanation for the coexistence of species in ecological communities. intuitively simple, but deceptively so. We show, via discussion and examples, that is not one mechanism of coexistence, but rather summarizes a set of similar.
As a result, there is a considerable body of research examining the effects of disturbance on species diversity, much of which focuses on testing the intermediate disturbance hypothesis (IDH).
The intermediate disturbance hypothesis is based on the following premises: Figure 18.2.1 18.2. 1: I describes how, at low levels of ecological disturbance species richness decreases as competitive exclusion increases; II shows that at intermediate levels of disturbance, diversity is maximized because species that thrive at both early and late ...
Defining and defending Connell's intermediate disturbance hypothesis: a response to Fox. In his recent Opinion article in TREE, Jeremy Fox [1] finds that evidence for the intermediate disturbance hypothesis (IDH) appears mixed and shows that some explanations linking disturbance to species coexistence are flawed.
Connell coined the term 'intermediate disturbance hypothesis' to describe this process. We strolled through the larger of his rainforest plots, avoiding stinging trees, biting flies, ticks and ...
Intermediate Disturbance Hypothesis. The intermediate disturbance hypothesis (IDH) suggests that local species diversity is maximized when ecological disturbances are neither too rare nor too frequent (Figure 17.1). At low levels of disturbance, more competitive organisms will push subordinate species to extinction and dominate the ecosystem.
A simple unimodal model with peak at 47.7% disturbance best explained vascular plant species richness in relation to human disturbance extent: species richness peaked at intermediate disturbance ...
In his recent Opinion article in TREE, Jeremy Fox [1] finds that evidence for the intermediate disturbance hypothesis (IDH) appears mixed and shows that some explanations linking disturbance to species coexistence are flawed. Based on this, he argues that we should abandon the IDH. Whereas we agree with his observations, we reject his conclusions. Fox's criticisms are misdirected because the ...
I appreciate Sheil and Burslem's thoughtful comments [1] on my opinion piece [2] arguing for abandonment of the intermediate disturbance hypothesis (IDH). I am gratified that they agree with my substantive criticisms of several ideas relating disturbance and species diversity. They argue, however, that those criticisms are directed at ideas that, although related to the IDH, are not actually ...
The intermediate disturbance hypothesis (IDH) suggests that the peak of species diversity occurs at intermediate-scale disturbances. The IDH received criticisms because many studies have shown that the relationship between disturbances and species diversity is generally not unimodal. We searched Web of Science for articles on IDH to study the applications of the hypothesis in animal and plant ...
In ecological disturbance: Intermediate disturbance hypothesis. Some ecologists claim that these qualitative traits—namely, the persistence of large disturbed patches and the relatively rapid recovery of smaller ones—may be synthesized through the intermediate disturbance hypothesis. This hypothesis states that a disturbance regime (or ...
PLATT, W.J., COLONIZATION AND FORMATION OF EQUILIBRIUM PLANT SPECIES ASSOCIATIONS ON BADGER DISTURBANCES IN A TALL-GRASS PRAIRIE, ECOLOGICAL MONOGRAPHS 45: 285 (1975). Google Scholar. RICKLEFS, R.E., ENVIRONMENTAL HETEROGENEITY AND PLANT SPECIES-DIVERSITY - HYPOTHESIS, AMERICAN NATURALIST 111: 376 (1977). Google Scholar.
The intermediate disturbance hypothesis predicts that the highest diversity will occur at levels of moderate disturbance. We measured the species diversity, abundance, and species diversity of bacteria on the human body in relation to disturbance by washing. Bacteria were sampled from three surfaces on the human body with varying disturbances ...
These disturbances don't impact the entire ecosystem, but just a section. Examples of intermediate disturbances in a temporal forest might be: lightening strikes and other small fires. blowdown of trees. river meander or flooding. landslides. small patches of logging. These occur occassionally throughout the landscape allow the ecosystem to: